|
全国免费电话:400-668-6834 -- 仅限中国地区 -- |
在线订购:QQ: 4006686834 电话订购:021-68591985 邮件订购:info@selleck.cn 我们也提供销售人员上门服务 |
技术支持电话:400-168-6834 我们将于一个工作日与您联系 |
生物描述
| 特异性 | Phospho-4E-BP1 (Thr37) Antibody [L9H8] 仅识别 Thr37 处磷酸化的内源性 4E-BP1 蛋白水平。 |
|---|---|
| 背景 | 磷酸化4E-BP1 (Thr37) 代表了帽依赖性翻译通路中eIF4E结合蛋白1的一种早期、mTOR依赖性的调控状态,它作为一种启动修饰,整合了来自PI3K-Akt-mTOR通路的生长因子和营养信号,从而实现多位点磷酸化并调控翻译起始。低磷酸化的4E-BP1通过位于包含Thr37和Thr46的中心区域附近的典型YXXXXLΦ基序与mRNA帽结合蛋白eIF4E紧密结合,阻碍eIF4G的对接,并阻止eIF4F复合物的组装。eIF4F复合物负责将40S核糖体亚基募集到5′帽上。同时,N端和C端区域稳定了这种抑制构象,并响应上游激酶的输入。当 4E-BP1 与 eIF4E 形成复合物时,FRAP/mTOR 直接在 Thr37 和 Thr46 位点磷酸化 4E-BP1,生成与体内两个最显著的 4E-BP1 磷酸肽共迁移的磷酸肽,并将 Thr37 和 Thr46 鉴定为 mTOR 的主要靶位点;即使在血清饥饿的细胞中,这些残基也高度磷酸化,并且存在于所有电泳不同的磷酸化 4E-BP1 亚型中,包括与 eIF4E 结合的形式。 Thr37 和 Thr46 的磷酸化不会破坏 4E-BP1-eIF4E 复合物或改变电泳迁移率,但突变分析表明,Thr37/Thr46 的磷酸化对于 C 端半段下游“血清敏感”位点(Thr70 和 Ser65)的有效后续磷酸化是必需的,因为 Thr37Ala、Thr46Ala 或双 Thr37Ala/Thr46Ala 取代会使 4E-BP1 的总磷酸掺入量减少 10-20 倍,并消除 C 端磷酸肽的出现,而不会损害基础 eIF4E 结合,这定义了磷酸化 Thr37/Thr46 是 eIF4E 分级磷酸化和释放的必要启动事件。在饥饿条件下,Thr37 和 Thr46 的磷酸化对雷帕霉素和 LY294002 敏感,并由 PI3K-Akt-mTOR 轴驱动:PI3K 或 Akt 抑制,或雷帕霉素敏感的 mTOR 阻断,均可降低 Thr37/Thr46 的磷酸化水平;而组成型活性 Akt 则促进血清刺激期间靶向的相同位点的磷酸化,这使得 Thr37 磷酸化的 4E-BP1 成为 Akt 下游 mTORC1 输出的近端报告分子。在果蝇中,同源物 d4E-BP 表现出 Thr37 和 Thr46 之间保守的调控相互作用,其中胰岛素刺激的 Thr46 磷酸化(可通过磷酸化 Thr37/46 抗体检测)是将 d4E-BP 从 eIF4E 结合的翻译抑制因子转化为结合力较弱形式的主要事件;这种调控依赖于胰岛素-PI3K-Akt-TSC-TOR信号通路,因为针对Dp110(PI3K)、dAkt、dTSC1或dTOR的RNAi分别会降低或增加d4E-BP Thr37/Thr46的磷酸化水平,并以符合线性Akt→TSC→TOR→4E-BP通路的方式改变d4E-BP的亚型模式。果蝇的层级分析表明,在血清饥饿的细胞中,d4E-BP在不阻止eIF4E相互作用的位点上携带多个磷酸基团。胰岛素以雷帕霉素敏感的方式触发Thr46位点的额外磷酸化,这与酸性更强的亚型的出现和eIF4E结合的减少相一致。这在功能上反映了哺乳动物4E-BP1中Thr37/Thr46磷酸化启动的必要性,该磷酸化启动后续修饰,从而释放eIF4E并允许eIF4F组装。这些发现表明,磷酸化 4E-BP1 (Thr37) 处于一个两步机制中,其中 mTORC1 首先磷酸化 eIF4E 结合的 4E-BP1 上的 Thr37/Thr46,以产生启动中间体,然后其他激酶——仍然在 mTORC1 信号传导环境中运行——修饰 Thr70 和 Ser65,最终导致 eIF4E 结合的丧失,从抑制性翻译状态转变为允许性翻译状态,以及生长和存活相关蛋白的合成增加。 |
使用信息
| 抗体应用 | WB | 稀释比例 |
|
||
|---|---|---|---|---|---|
| 反应性 | Human | ||||
| 抗体类型 | Rabbit Monoclonal Antibody | MW | 13 kDa | ||
| 储存液配方 | PBS, pH 7.2+50% Glycerol+0.05% BSA+0.01% NaN3 | 储存条件 (自收到货起) |
-20°C (avoid freeze-thaw cycles), 2 years | ||
文献参考
|
Application Data
WB
Validated by Selleck
-
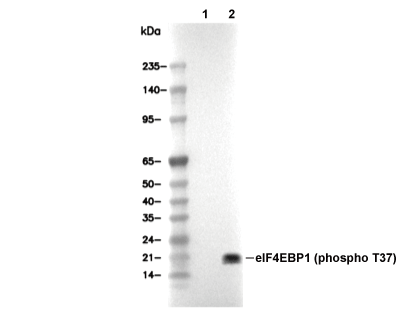 Lane 1: 293, Lane 2: 293 (insulin treated)
Lane 1: 293, Lane 2: 293 (insulin treated)