|
全国免费电话:400-668-6834 -- 仅限中国地区 -- |
在线订购:QQ: 4006686834 电话订购:021-68591985 邮件订购:info@selleck.cn 我们也提供销售人员上门服务 |
技术支持电话:400-168-6834 我们将于一个工作日与您联系 |
生物描述
| 特异性 | Phospho-PLCβ3 (Ser537) Antibody [K10E11] 仅识别 Ser537 处磷酸化的内源性 PLCβ3 蛋白水平。 |
|---|---|
| 背景 | 磷酸化 PLCβ3 (Ser537) 代表磷脂酰肌醇特异性 PLC 家族中磷脂酶 C β 3 的一个具有重要功能意义的调节状态,它将 G 蛋白偶联和受体酪氨酸激酶信号传导与磷脂酰肌醇 4,5-二磷酸水解和下游钙-PKC 通路联系起来,PLCβ3 可催化肌醇 1,4,5-三磷酸和二酰甘油的生成,以响应包括 VEGF、凝血酶和其他 GPCR 激动剂在内的各种细胞外信号。 PLCβ3 具有一个 N 端 pleckstrin 同源结构域,其后依次为 EF-hand 结构域、紧密结合的 X-Y 催化核心(由一个短连接肽分隔)以及 C 端 C2 结构域。这些结构域共同协调膜结合、钙结合和底物结合,而 C 端调控区则整合来自 Gαq/11、Gβγ 和蛋白激酶的信号。Ser537 位于 PLCβ3 N 端半段,在 X 结构域上游,在内皮细胞中经 VEGFR2 刺激后迅速磷酸化,30 秒内即可检测到磷酸化,1 分钟内达到最大水平,同时抑制性 Ser1105 位点的磷酸化以及 PLCγ1 在 Tyr783 位点的酪氨酸磷酸化也同时发生。 VEGF激活VEGFR2可同时激活经典的PLCγ1和PLCβ3通路。VEGFR2选择性激酶抑制剂可消除VEGF诱导的PLCβ3在Ser537和Ser1105位点的磷酸化,表明Ser537修饰位于VEGF-VEGFR2受体酪氨酸激酶级联反应的下游,而非一般的GPCR信号通路。PLCβ3 Ser537磷酸化伴随催化性PIP2水解的增强,表现为VEGF刺激的内皮细胞中IP1/IP3生成增加。PLCβ3敲低可降低VEGF诱导的肌醇磷酸积累,表明PLCβ3与PLCγ1共同直接参与VEGF驱动的磷脂酰肌醇周转。 VEGF-VEGFR2-PLCβ3信号通路通过将PIP2水解和二酰甘油生成与小GTP酶CDC42的激活偶联,从而调控内皮细胞的定向迁移。PLCβ3的缺失会选择性地抑制CDC42的激活,而RhoA和Rac1的反应则不受影响。这与定向迁移的丧失、应力纤维形成减少以及血管生成过程中芽尖丝状伪足相关导向结构的减少相关。在胚状体血管生成模型中,VEGF刺激可诱导萌芽内皮细胞中PLCβ3的募集和磷酸化,磷酸化的PLCβ3定位于CD31阳性的血管结构,表明包括Ser537磷酸化部分在内的磷酸化PLCβ3在血管生成前沿信号传导和血管萌芽中发挥作用。 PLCβ3 是内皮细胞增殖的负调控因子,因为敲低 PLCβ3 会加速细胞周期进程,增加 S 期比例,提高细胞周期蛋白 D1 和 CDC2 的表达,而不会改变 VEGF 诱导的 ERK、JNK 或 p38 激活,这使得 Ser537 修饰的 PLCβ3 处于血管生成过程中平衡增殖与迁移的信号分支中。 PLCβ3 上的 Ser537 和 Ser1105 残基在 VEGF 和 GPCR 激动剂(如凝血酶和 PAR 配体)的作用下均发生磷酸化,这使得 Ser537 处于一个更广泛的磷酸化网络中,其中 PKA 或 PKC 对 Ser1105 的磷酸化抑制了 PLCβ3 的酶活性,而 CaMKII 对 Ser537 的磷酸化则有助于基础或刺激调节活性,因此磷酸化 PLCβ3 (Ser537) 可作为生长因子和 GPCR 通路中 Ca2+/CaMKII 连接的 PLCβ3 调节的读数。 VEGF–VEGFR2–PLCβ3 轴与 Gq/11 和 Gβγ 亚基相互作用,因为 VEGFR2 激活会激活内皮细胞中 PLCβ3 上游的 G 蛋白信号传导,而 PLCβ3 则在磷脂酰肌醇水解和 CDC42 依赖性细胞骨架重塑的水平上整合 RTK 和 GPCR 输入。 |
使用信息
| 抗体应用 | WB | 稀释比例 |
|
||
|---|---|---|---|---|---|
| 反应性 | Human, Mouse, Rat | ||||
| 抗体类型 | Rabbit Monoclonal Antibody | MW | 150 kDa | ||
| 储存液配方 | PBS, pH 7.2+50% Glycerol+0.05% BSA+0.01% NaN3 | 储存条件 (自收到货起) |
-20°C (avoid freeze-thaw cycles), 2 years | ||
文献参考
|
Application Data
WB
Validated by Selleck
-
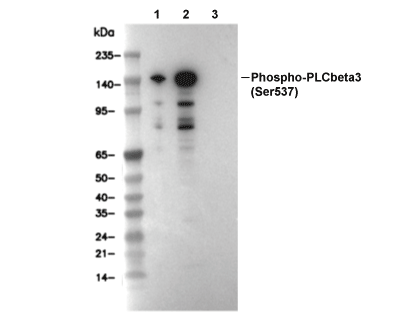 Lane 1: 293T, Lane 2: 293T (Anisomycin, 25 ug/ml, 30 min), Lane 3: 293T (Anisomycin, 25 ug/ml, 15 min; λ phosphatase treated)
Lane 1: 293T, Lane 2: 293T (Anisomycin, 25 ug/ml, 30 min), Lane 3: 293T (Anisomycin, 25 ug/ml, 15 min; λ phosphatase treated)