|
全国免费电话:400-668-6834 -- 仅限中国地区 -- |
在线订购:QQ: 4006686834 电话订购:021-68591985 邮件订购:info@selleck.cn 我们也提供销售人员上门服务 |
技术支持电话:400-168-6834 我们将于一个工作日与您联系 |
生物描述
| 特异性 | VEGFA C-terminal Antibody [D1C9] 可检测内源性 VEGFA 总蛋白水平。 |
|---|---|
| 背景 | 血管内皮生长因子A (VEGFA) 的C端结构域通过外显子5、7、8a和8b的选择性剪接决定其亚型特异性,产生的变体对VEGF受体2 (VEGFR2) 的亲和力相似,但在与神经纤毛蛋白-1 (NRP1) 和硫酸乙酰肝素蛋白聚糖 (HSPG) 的结合方面存在显著差异。NRP1和HSPG作为重要的辅助受体,调节受体的呈递和信号强度。由外显子5和7编码的C端区域包含一个肝素结合结构域 (HBD),该结构域具有结构灵活性,并包含两个不同的碱性氨基酸簇——R13、R14、K15残基和K30、R35、R49残基——它们通过静电相互作用协同结合肝素和硫酸乙酰肝素。这种结构域的灵活性使得VEGFA能够发生动态构象变化,而这种变化对于肝素结合至关重要。C端残基S34、C48和D51在毫秒级的时间尺度上发生运动,并在与肝素八糖结合后趋于稳定,从而促进受体复合物的组装和在细胞外基质中的定位。包含由外显子8a编码的六个氨基酸的VEGFA亚型与来自外显子5或7的结构域协同作用,以实现高效的NRP1和HSPG结合。而包含外显子8b的变体则表现出不同的功能特性——VEGF165b与VEGFR2结合,但不触发下游磷酸化级联反应,并作为血管生成抑制剂发挥作用;而最短的外显子8b变体VEGF111b却出人意料地表现出强大的促血管生成活性。这表明,结构域组成通过超越简单受体亲和力的机制决定了功能结果。 C端结构域通过与硫酸肝素蛋白聚糖(HSPG)相互作用控制VEGFA在细胞外基质中的滞留,从而调节VEGFA的生物利用度;并通过募集NRP1来调节受体激活动力学,NRP1可增强VEGFR2信号传导,并通过激活PI3K/AKT和MAPK/ERK通路促进内皮细胞的迁移、增殖和存活。C端结构域启动的NRP1结合使VEGFA的功能超越内皮细胞,通过涉及信号素通路组分的不同信号网络介导轴突导向和神经元迁移。C端氢键结合域(HBD)固有的结构灵活性使其能够与多种结合伙伴相互作用,同时保持特异性,从而使VEGFA能够协调多种细胞反应,包括血管通透性调节、基底膜重塑以及与缺氧诱导因子信号网络的整合。可变剪接产生不同的C端结构,从而形成一系列具有不同血管生成能力的VEGFA亚型——基质结合的较长亚型建立局部信号梯度,引导血管萌芽;而可自由扩散的较短亚型则促进远端内皮细胞活化,共同调控血管模式形成和成熟。C端结构域表达模式的失调会导致病理性血管生成,肿瘤细胞选择性地产生外显子8a亚型,以最大程度地增强NRP1介导的信号传导和血管通透性,从而促进营养物质输送和转移扩散。而通过抗VEGF抗体或可溶性诱饵受体靶向C端结构域功能的治疗策略则会破坏VEGFR2和NRP1的结合。 |
使用信息
| 抗体应用 | IHC, IF, FCM | 稀释比例 |
|
||||||
|---|---|---|---|---|---|---|---|---|---|
| 反应性 | Mouse, Human | ||||||||
| 抗体类型 | Rabbit Monoclonal Antibody | MW | 44 kDa | ||||||
| 储存液配方 | PBS, pH 7.2+50% Glycerol+0.05% BSA+0.01% NaN3 | 储存条件 (自收到货起) |
-20°C (avoid freeze-thaw cycles), 2 years | ||||||
文献参考
|
Application Data
IHC
Validated by Selleck
-
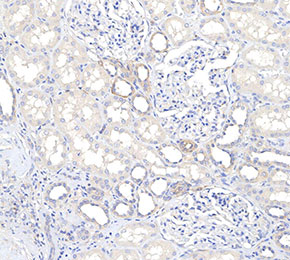 Immunohistochemical analysis of formalin fixed paraffin embedded human kidney tissue with F0064 at 1:100 dilution.
Immunohistochemical analysis of formalin fixed paraffin embedded human kidney tissue with F0064 at 1:100 dilution.