IFN-γ Antibody [A12D6]
目录号: F6371
抗体应用:
反应性:
-
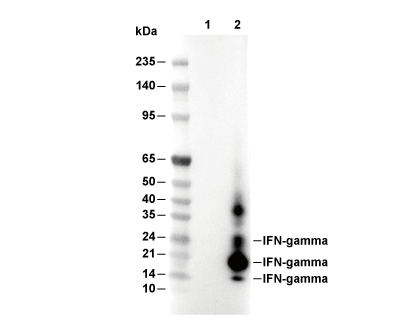 Lane 1: 293T (mock transfected), Lane 2: 293T (mIFN-γ transfected)
Lane 1: 293T (mock transfected), Lane 2: 293T (mIFN-γ transfected)
使用信息
| 稀释比例 |
|---|
|
| 抗体应用 |
|---|
| WB |
| 反应性 |
|---|
| Mouse |
| 抗体类型 |
|---|
| Rabbit Monoclonal Antibody |
| 储存液配方 |
|---|
| PBS, pH 7.2+50% Glycerol+0.05% BSA+0.01% NaN3 |
| 储存条件(自收到货起) |
|---|
| -20°C (avoid freeze-thaw cycles), 2 years |
|
预测分子量
实际分子量
|
|---|
|
18 kDa
17 kDa, 19 kDa, 23 kDa
|
| *为什么预测分子量和实际分子量会有不同? 以下原因可能会导致蛋白分子量与预测不同:翻译后修饰(磷酸化/糖基化);剪接变体和异构体;相对电荷;多聚体。 |
生物描述
| 特异性 |
|---|
| IFN-γ Antibody [A12D6] 可检测内源性 IFN-γ 总蛋白水平。 |
| 克隆号 |
|---|
| A12D6 |
| 别名 |
|---|
| gamma interferon; Ifg; IFN-g; IFN-gamma; Ifng; Interferon gamma; interferon, gamma |
| 背景 |
|---|
| 干扰素-γ (IFN-γ) 是唯一的 II 型干扰素,它是一种多效性细胞因子,通过作用于除成熟红细胞以外的几乎所有细胞类型,协调先天性和适应性免疫反应。IFN-γ 以非共价连接的同源二聚体形式存在,由两条反平行排列的多肽链组成,每条多肽链包含六个 α 螺旋,形成具有二重对称性的紧密球状结构。其生物活性需要氨基末端残基 1-10 和羧基末端残基 129-143 才能完全结合受体。该蛋白发生 N-糖基化修饰,形成成熟的亚基,在 SDS-PAGE 电泳中迁移至约 25 kDa 处。然而,糖基化修饰的作用是保护蛋白免受蛋白水解酶的攻击,而非直接发挥功能活性。 IFN-γ 的产生主要局限于 T 淋巴细胞(Th1 CD4+ 和细胞毒性 CD8+)和自然杀伤细胞,NK 细胞保持组成型 IFNG 位点可及性,能够在感染或癌症期间通过受体介导的激活快速分泌 IFN-γ。这种激活是通过免疫受体酪氨酸激活基序触发 Src/MAPK/ERK/p38 通路,激活 Fos 和 Jun 转录因子;此外,细胞因子介导的激活主要通过 IL-12 与 IL-12 受体结合,激活 STAT4 和 NF-κB,建立有利于 Th1 反应的正反馈回路。该蛋白结合由IFNGR1(90 kDa,主要配体结合剂)和IFNGR2(62 kDa,亲和力增强剂)组成的异二聚体受体复合物。化学计量比表明,每个IFN-γ同源二聚体结合两个IFNGR1亚基和两个IFNGR2亚基,形成一个2:2:2的信号传导活性复合物。IFNGR1在膜近端LPKS基序处具有组成型JAK1结合位点,其中Pro267对功能至关重要;此外,在YDKPH基序处还有一个可诱导的STAT1对接位点,Tyr440的磷酸化可启动STAT1 SH2结构域的募集。IFNGR2在PPSIPLQIEEYL序列处具有组成型JAK2结合位点。 IFN-γ 配体结合诱导 IFNGR1-IFNGR2 异二聚化,使组成型受体结合但处于非活性状态的 JAK1 和 JAK2 彼此靠近,从而实现自身磷酸化和反式磷酸化,激活这两个激酶。激活的 JAK 磷酸化 IFNGR1 的 Tyr440 位点,形成 STAT1 的结合位点。募集的 STAT1 在此位点经历 JAK 介导的 Tyr701 位点磷酸化以及丝氨酸激酶介导的 Ser727 位点磷酸化,最终被完全激活。磷酸化的 STAT1 分子通过 SH2 结构域与磷酸酪氨酸的相互作用形成同二聚体,从受体解离,转位至细胞核,并与具有 TTNCNNNAA 共有序列的 γ 激活位点 (GAS) DNA 元件结合,驱动干扰素刺激基因的转录,包括转录因子 IRF1、IRF2、IRF8、IRF9 和 RELA,这些转录因子随后激活第二波 ISG。 IFN-γ 信号传导受到以下因素的负调控:含有 SH2 结构域的磷酸酶(SHP 蛋白)与 IFNGR 复合物组成性结合,使激活的酪氨酸残基去磷酸化;细胞因子信号抑制因子(SOCS)蛋白由 IFN-γ 转录诱导,抑制 JAK 并使其泛素化以便降解;以及活化 STAT 蛋白抑制剂(PIAS)阻止 STAT1 与 DNA 的相互作用。T 细胞蛋白酪氨酸磷酸酶 TCP45 使核内 STAT1 去磷酸化,从而实现细胞质循环。 IFN-γ 具有多种功能,包括激活巨噬细胞(作为主要的巨噬细胞激活因子),诱导诱导型一氧化氮合酶的表达,从而产生杀菌性一氧化氮;产生促炎细胞因子 IL-1β、TNF、IL-12、IL-18 和 IL-23;上调 MHC I 类和 II 类分子以及共刺激蛋白 CD80 和 CD86,从而增强抗原呈递;诱导免疫蛋白酶体成分 LMP1、LMP7 和 MECL1,从而改变肽库;上调 TAP-1 和 TAP-2 转运蛋白,将肽转运到内质网;以及激活 MHC II 类转录激活因子 CIITA,从而驱动 II 类表达。该细胞因子通过 STAT1 激活 T-bet 转录因子促进 Th1 分化,上调 IL-12 受体和 IFN-γ 表达,建立正反馈;同时通过维持 IFNGR 表达抑制 Th2 和 Th17 增殖,使这些亚群对 IFN-γ 敏感;促进 B 细胞免疫球蛋白类别从 IgE 向 IgG2a 转换,促进抗体依赖性细胞毒性;通过上调 IL-2 受体、T-bet 和颗粒酶表达增强 CD8+ T 细胞的细胞溶解能力;并激活 NK 细胞对肿瘤细胞和受感染靶细胞的细胞毒性。 IFN-γ在癌症免疫编辑中发挥关键作用,免疫选择可清除表达强新抗原的肿瘤细胞。在IFN-γ不敏感的IFNGR1缺陷或STAT1缺陷小鼠中,肿瘤的发生频率和速度均高于野生型小鼠。免疫介导的清除需要宿主细胞和肿瘤细胞中的IFN-γ信号通路。实验表明,IFN-γ不敏感的肿瘤会持续生长,而IFN-γ敏感的肿瘤在脂多糖治疗后会被清除。然而,该蛋白也具有促肿瘤作用,例如通过上调肿瘤细胞和免疫细胞上的程序性死亡配体1 (PD-L1) 和PD-L2,增强适应性免疫抵抗;通过上调促炎细胞因子和Th17反应促进乳头状瘤的发生;以及通过不受控制的IFN-γ信号通路促进SOCS1缺陷小鼠发生结直肠癌。 IFN-γ除了激活JAK-STAT通路外,还能激活其他信号通路,包括PI3K/Akt/mTOR/p70S6激酶轴(该轴是干扰素刺激基因翻译所必需的,从而产生抗病毒作用)、MAPK通路Pyk2和ERK1/2、TGFβ/SMAD信号通路(该通路诱导NADPH氧化酶NOX1和NOX4产生活性氧,进而引发DNA损伤和细胞衰老),以及不依赖于STAT1的C/EBPβ和巨噬细胞炎症蛋白MIP-1α和MIP-1β的转录。该细胞因子调节造血功能,其过表达可导致骨髓抑制,进而引发骨髓增生异常综合征和再生障碍性贫血;它还具有抗血管生成作用,通过激活ERK1/2通路参与神经发生和神经元分化,并通过mTORC1和MNK激酶调控巨噬细胞代谢,这些激酶最终汇聚于翻译起始因子eIF4E。 |
| 参考文献 |
|---|
|
技术支持
在订购、运输、储存和使用我们的产品的任何阶段,您遇到的任何问题,均可以通过拨打我们的热线电话400-668-6834,或者技术支持邮箱tech@selleck.cn,直接联系到我们。我们会在24小时内尽快联系您。
* 必填项


